SCFAs made their first major debut in 2009, when they were shown to protect against colitis (3). Two clues led to this breakthrough: i) germ-free mice (which lack microbes and thus, microbial-derived metabolites) displayed exacerbated colitis, ii) mice that had a normal microbiota but lacked a G-protein coupled receptor 43 (Gpr43) showed a similar trend. The researchers linked these two observations and hypothesized that microbial products (clue #1) interact with GPR43 (clue #2), resulting in the mitigation of colitis. Knowing that SCFAs bind GPR43 (4), the researchers administered acetate to both wild-type (WT) and Gpr43-/- mice, followed by the experimental induction of colitis, which was when the breakthrough occurred: Gpr43-/- mice had exacerbated symptoms, which proved that acetate reduces inflammation via GPR43 (3). Since then, many others have expanded on this observation, noticing that bifidobacteria-derived acetate induced anti-inflammatory gene expression in colonocytes, increased transepithelial resistance and protected mice against enteropathogenic infection (5). In 2013, three independent studies pointed out an entirely new function of SCFAs, which today is considered their hallmark feature: the ability to induce regulatory T cell (Treg) differentiation (6-8) (Fig.1).
 It is important to note that the described mechanism is not exclusive. Several other groups showed the capacity of SCFAs to mediate anti-inflammatory effects in Treg-independent ways, e.g. via modulation of a dendritic cell function (9), alteration of hematopoiesis in the bone marrow (10), or activation of inflammasome (11).
Although these observations were fascinating on their own, what truly inspired us to share this story were the recent findings from the aforementioned human study by Duscha et al.
It is important to note that the described mechanism is not exclusive. Several other groups showed the capacity of SCFAs to mediate anti-inflammatory effects in Treg-independent ways, e.g. via modulation of a dendritic cell function (9), alteration of hematopoiesis in the bone marrow (10), or activation of inflammasome (11).
Although these observations were fascinating on their own, what truly inspired us to share this story were the recent findings from the aforementioned human study by Duscha et al.

SCFAs induce Treg differentiation
Arpaia and his colleagues started with a simple question: are SCFAs acting directly on naive CD4+ T cells? To answer it, they differentiated naive CD4+ T cells into a Treg subset in vitro, in the presence of increasing concentrations of butyrate. Interestingly, this led to a dose-dependent increase in the frequency of differentiated Tregs, along with the stabilization of FoxP3, their master transcription factor (6). This finding was confirmed and extended by Furusawa et al., who identified detailed epigenetic signatures linked to FoxP3 stabilization. They noted histone H3 acetylation at the promoter site and at the intragenic enhancer element 3 of Foxp3 gene locus, occurring one day prior to Foxp3 expression (7). Finally, Smith and his colleagues recapitulated these observations using propionate, which inhibited histone deacetylation (thus, increased histone acetylation), and contributed to a higher expression of FoxP3 in colonic Tregs. Importantly, those Tregs were more efficient at suppressing effector T cell responses than Tregs from water-treated controls (8).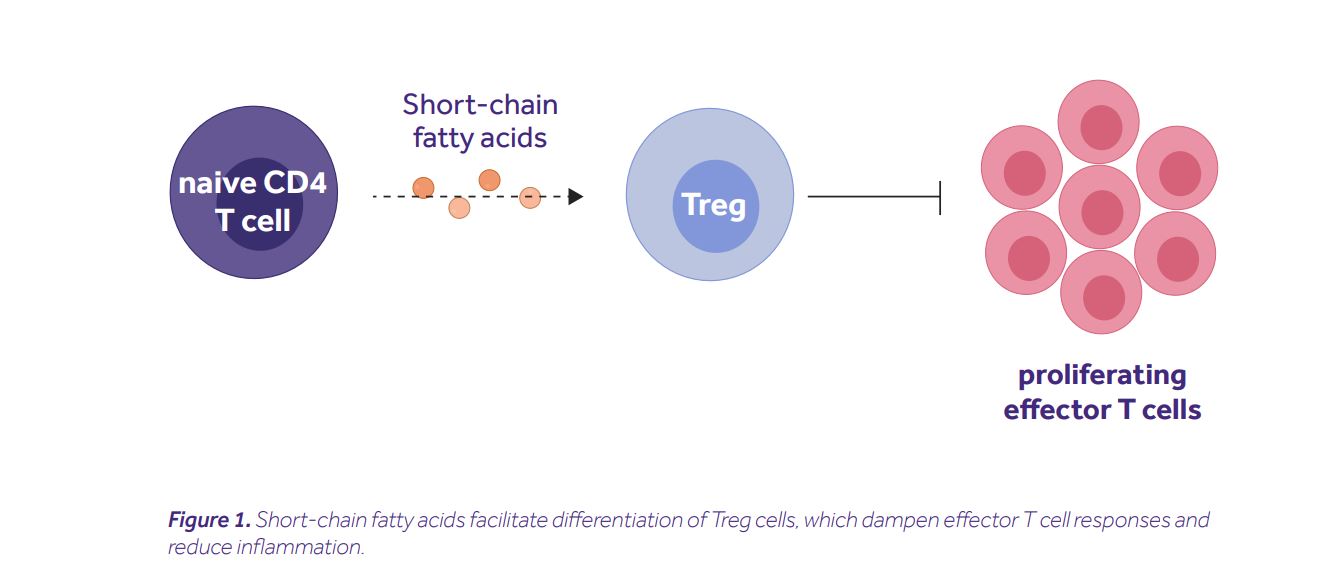 It is important to note that the described mechanism is not exclusive. Several other groups showed the capacity of SCFAs to mediate anti-inflammatory effects in Treg-independent ways, e.g. via modulation of a dendritic cell function (9), alteration of hematopoiesis in the bone marrow (10), or activation of inflammasome (11).
Although these observations were fascinating on their own, what truly inspired us to share this story were the recent findings from the aforementioned human study by Duscha et al.
It is important to note that the described mechanism is not exclusive. Several other groups showed the capacity of SCFAs to mediate anti-inflammatory effects in Treg-independent ways, e.g. via modulation of a dendritic cell function (9), alteration of hematopoiesis in the bone marrow (10), or activation of inflammasome (11).
Although these observations were fascinating on their own, what truly inspired us to share this story were the recent findings from the aforementioned human study by Duscha et al.
SCFAs in multiple sclerosis patients
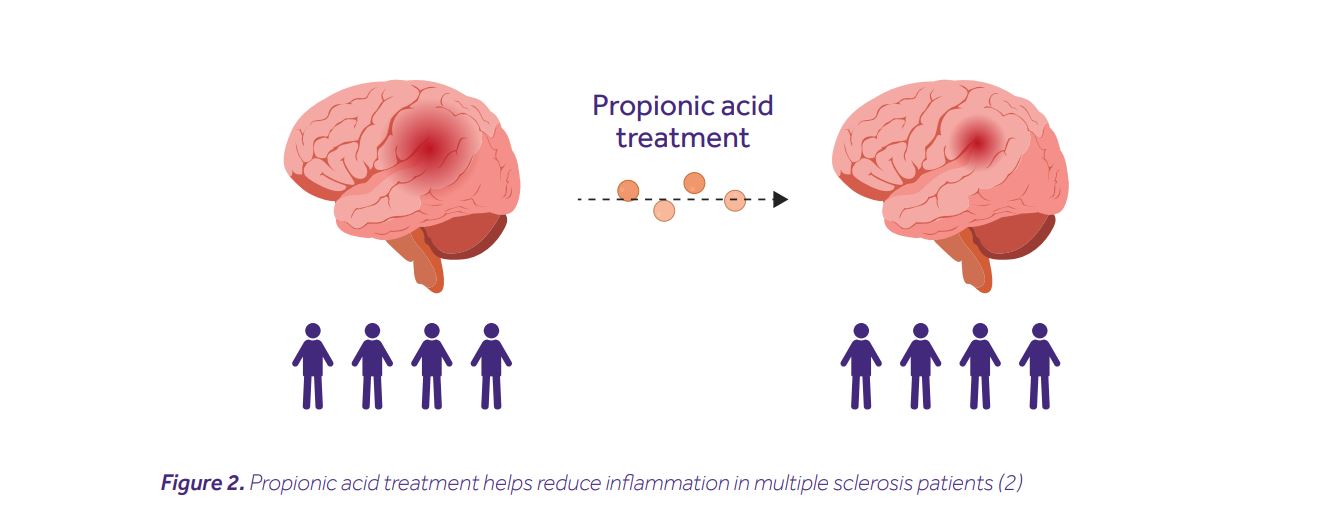
It all started with a simple question: do SCFA levels vary between healthy individuals and MS patients? In the quest for an answer, Duscha and his colleagues recruited two multiple sclerosis cohorts (made up of 268 and 36 patients respectively), and a healthy control cohort (of 68 individuals). They then measured their SCFA concentrations from serum and stool samples. Although no differences in the levels of butyric or acetic acids were noted, MS patients from both cohorts had significantly lower levels of propionic acid (PA). This was accompanied by differences in their microbiota composition, e.g. depletion of SCFA-producing Butyricimonas, and an increase of Flavonifractor, Escherichia, Shigella and Collinsella. MS patients are known to have imbalanced T helper cell responses, i.e. increased frequencies of pathogenic Th17 cells and decreased frequencies of anti-inflammatory Tregs. This gave the researchers the rationale to hypothesize that PA treatment may restore this imbalance and improve a disease course. Indeed, PA treatment increased the frequency of Tregs in the blood, restored their mitochondrial respiration, elevated secretion of interleukin-10, and collectively improved the capacity to suppress effector immune responses. Most remarkably, however, patients receiving a prolonged PA treatment (>1 year) had a reduced annual relapse rate, along with a lower risk of disease progression and brain atrophy (Fig. 2).